1. INTRODUCTION
Stroke is the third leading cause of death in the United States and results in substantial health-
care expenditures; the mean lifetime cost resulting from an ischemic stroke is estimated at
$140,048 per patient, and this estimation is higher for people over 45 years. Nationwide in 2010,
the estimated direct and indirect costs of stroke totaled $73.7 billion [1]. Although many clinical
trials have been completed in stroke patients, none of these have demonstrated protective
efficacy except for thrombolysis [2, 3]. In the case of cardiac arrest and resuscitation only
hypothermia has been shown to have clinical utility [4]. In some sense the two therapies that
have been effective thus far clinically have broad targets, and do not only affect a single injury
mechanism. In contrast, of the failed trials, many targeted neuron-specific injury mechanisms
[5]. This may reflect too narrow a view of what is needed for brain preservation. A large body of
work has shown that astrocytes play key roles both in normal and pathological central nervous
system functioning [6]. Astrocytes are the most abundant brain cell type, and in addition to their
multiple important homeostatic roles, they organize the structural architecture of the brain, help
organize communication pathways, and modulate neuronal plasticity (for recent review see
[7, 8]). Thus, astrocytes are now thought to be important potential targets for manipulation.
Ischemic stroke is caused by an interruption of cerebral blood flow that leads to stress, cell death,
and inflammation. Neurons are more susceptible to injury than astrocytes when studied under
some in vitroconditions [9, 10]. Neurons have less endogenous antioxidants and are susceptible
to excito-toxicity [10]. Both normally and after ischemia, astrocytes support neurons by
providing antioxidant protection [11, 12], substrates for neuronal metabolism [13], and glutamate
clearance REF. Although astrocytes are sometimes more resilient than neurons, injury can result
in impaired astrocyte function even when astrocytes do not die. Impaired astrocyte function can
amplify neuronal death [14]. Therefore, many recent efforts have focused on the astrocyte-
neuron interaction and how astrocyte function can be improved after stroke to enhance neuronal
support and survival [10, 15, 16]. A growing body of data demonstrates that astrocytes play a
multifaceted and complex role in the response to ischemia, with potential to both enhance and
impair neuronal survival and regeneration [17]. Many recent studies focus on the astrocyte-
neuron interaction and several investigate ways in which astrocyte function can be improved
after stroke to enhance neuronal survival.
This review provides a brief overview of the pathophysiological events underlying ischemic
brain damage, and considers how these events affect astrocyte-mediated support of neurons. In
addition, we discuss some experimental approaches to enhance the neuronal supportive role of
astrocytes as a novel strategy against stroke. Finally, we explore how these approaches may
eventually be applied in the clinical setting to improve stroke outcome for patients.
2. ASTROCYTE VIABILITY AFTER ISCHEMIA
2.1. In Vitro Studies
In vitro studies have provided substantial insight into the mechanisms governing the survival of
astrocytes following simulated ischemia. These investigations have shown that astrocytes are
generally more resistant than neurons to oxygen-glucose deprivation (OGD) performed in media
at physiologically normal pH, an in vitro model of ischemia [10, 18]. Most neurons in astrocyte-
neuronal co-cultures will die after 60–90 min of OGD, while astrocyte cultures only suffer a
similar extent of injury after 4–6 hours [9, 18, 19]. Different astrocyte populations exist and
astrocytes isolated from different brain regions such as cortex, striatum, and hippocampus differ
in their sensitivity to OGD [15, 20, 21]. Furthermore, Lukaszevicz and colleagues [22] reported
that protoplasmic astrocytes lose their integrity faster than fibrous astrocytes, which may explain
the regional differences in susceptibility to ischemia between white matter astrocytes which are
fibrous and grey matter astrocytes that are protoplasmic. Although less susceptible to OGD-
induced damaged in vitro studies have highlighted certain elements that are highly toxic to
astrocytes. For example, acidosis has been found to be very effective in killing astrocytes
[23–26], in contrast to neurons, which are protected in acidic conditions [24, 26].
2.2. Focal Cerebral Ischemia
Much of the information about the recovery of astrocytes in vivo has been provided by studies
using immunohistological markers for astrocyte specific proteins, such as glial fibrillary acidic
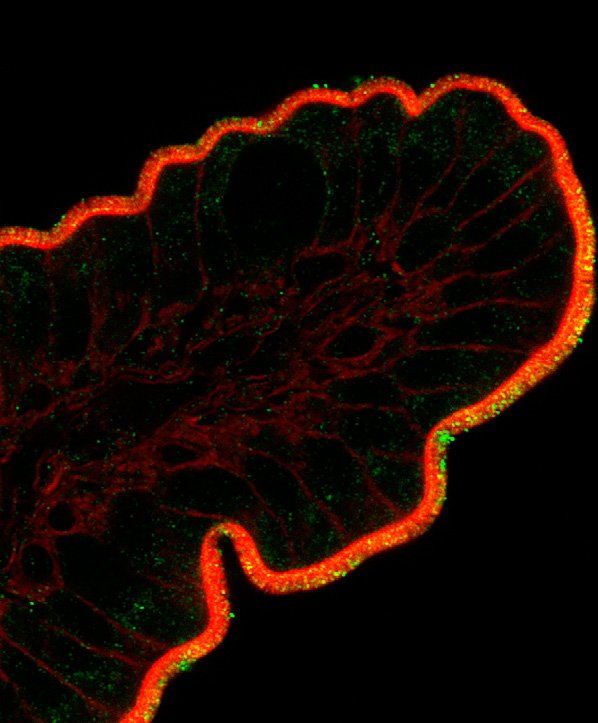
protein (GFAP) and glutamine synthetase GS; Fig. 1. Using these markers as tools, several
investigations suggest that astrocytes are better preserved than neurons in animal models of
stroke outside the core where all cells die [27–29]. Though neuronal markers are decreased as
soon as 1 hour after MCAO, GFAP expression is preserved over the first 3 hours of reperfusion
after 2 hour MCAO [29] and GS is increased 3 hours following a 3 hour MCAO [28]. At later
reperfusion periods, GFAP increases in the peri-infarct area that later develops into the glial scar
[29–32]. In contrast, Liu and colleagues [33] reported the deterioration of some astrocyte
markers prior to that of neuronal markers. Discrepancy in findings may be due to differences in
detection (i.e., protein vs. mRNA) and injury paradigms.
Fig. (1)
Expression of different astrocytic proteins following stroke. Increased expression of GFAP is a
hallmark of astrocytes activation, as is induction/re-expression of vimentin. Astrocytes normally
express glutamine synthetase (GS) and S100β, genes …
2.3. Forebrain Ischemia
Excitotoxic neuronal injury is a common mechanism in both acute and chronic
neurodegenerative diseases. It has long been appreciated that inhibition of astrocyte glutamate
uptake [34, 35], and more recently inhibition of astrocyte mitochondrial function [36], impairs
neuronal survival from excitotoxic injury. Brief forebrain ischemia is a model of the delayed
hippocampal neuronal loss seen in patients following cardiac arrest and resuscitation, and in part
involves excitotoxicity. Increased generation of reactive oxygen species (ROS) and
mitochondrial dysfunction in CA1 astrocytes contributes to ischemia-induced loss of GLT-1 and
ultimately to delayed death of CA1 neurons [15]. Our studies and those of other laboratories
have demonstrated that selective dysfunction of hippocampal CA1 subregion astrocytes, with
loss of glutamate transport activity and immunoreactivity for glutamate transporter 1 (GLT-1),
occurs at early reperfusion times, hours to days before the death of CA1 neurons [15, 37, 38].
The heterogeneous degeneration of astrocytic processes and mitochondria was tightly associated
with the appearance of disseminated selective neuronal necrosis and its maturation after
temporary ischemia [39]. By electronmicroscopy the same investigators [40] found that focal
infarction is exacerbated by temporary microvascular obstruction due to compression by swollen
astrocytic end-feet. However, hypoxia has multiple effects on astrocytes and their ability to
support neuronal viability [41]. For example, hypoxia induces astrocyte-dependent protection of
neurons following hypoxic preconditioning. Yet, hypoxia induces processes in astrocytes that
augment neuronal death in other situations, such as the coincidence of hypoxia with
inflammatory signaling.
3. REACTIVE ASTROGLIA: GOOD OR BAD AFTER STROKE?
The astrocyte response to ischemia has traditionally been viewed as detrimental to recovery, as
the astrocyte-rich glial scar has both physical and chemical inhibitory properties [42, 43]. As
components of the glial scar, astrocytes exhibit hypertrophied, interdigitated processes that form
a physical barrier. Astrocytes produce inhibitory molecules including chondroitin sulfate
proteoglycans (CSPGs) that contribute to chemical inhibition [44, 45]. In the acute setting,
astrocytic gap junctions may remain open following ischemia [46], allowing substances such as
proapoptotic factors to spread through the syncytium, thereby expanding the size of the infarct
[47]. As discussed below, astrocytes can also produce a variety of pro-inflammatory cytokines.
Many studies have shown that decreased astrogliosis often correlates with decreased infarct size.
Nonspecific inhibition of cell proliferation following ischemia using a cyclin kinase inhibitor
decreases astrocyte proliferation and results in improved functional recovery [48]. In addition,
treatment with alpha-melanocyte stimulating hormone [49], cysteinyl leukotriene receptor
antagonist [50], cliostazol [51], and caffeic acid [52] result in reduced infarct size accompanied
by a decrease in astrogliosis. Treadmill exercise [28] and acupuncture [53] are similarly
associated with improved outcome and reduced astrogliosis. Thus, results from several studies
suggest that treatments that decrease infarct size are often accompanied by attenuated astrocyte
response. Despite the frequent association of decreased astrogliosis with improved outcome, it is
difficult to determine cause and effect, since the extent of astrogliosis likely reflects the severity
of the injury, as well as influencing it.
In addition to their role in glial scar formation, astrocytes also respond to ischemia with functions
important for neuroprotection and repair. These include protecting spared tissue from further
damage [14], taking up excess glutamate as discussed above, rebuilding the blood brain barrier
[54, 55], and producing neurotrophic factors [10]. GFAP knockout mice exhibit larger lesions
than their wild-type littermates following focal ischemia [56], and mice lacking both GFAP and
vimentin have impaired astrocyte activation, decreased glutamate uptake abilities, and attenuated
PAI-1 expression after ischemia [57]. Application of astrocyte-conditioned media after transient
MCAO results in decreased infarct volume and regained blood-brain barrier function [58],
suggesting that factors released by astrocytes following ischemia are important for
neuroprotection.
Although few studies other than the use of animals lacking vimentin and GFAP have specifically
targeted astrocyte activation after ischemia, there is correlational evidence suggesting that
astrogliosis may be beneficial. Environmental enrichment, which results in reduced infarct size
and improved recovery following ischemia, also leads to increased astrocyte proliferation
[59, 60]. After focal ischemia, aged rats exhibit increased tissue damage and increased astrocyte
hypertrophy, but have decreased astrocyte proliferation compared to young rats [61]. Systemic
infusion of bone marrow stromal cells following MCAO increases gliogenesis and decreases
lesion size [62, 63]. In addition, administration of transforming growth factor α (TGFα), a known
mitogen for astrocytes [64], following MCAO leads to reduced infarct size and improved
functional recovery [65]. Furthermore, ischemic preconditioning that produces a neuroprotective
state leads to prolonged astrocyte expression of Hsp27 [66]. Finally, mice lacking connexin 43,
the gap junction connecting astrocyte networks that is needed for proper neurotransmitter and
potassium regulation, have increased infarcts following MCAO [67]. Thus, astrocytes have the
potential to be both detrimental and beneficial following ischemic insult, making them promising
targets for manipulation to improve outcome.
4. ASTROCYTE-MEDIATED INFLAMMATION AFTER STROKE: A DOUBLE-EDGED
SWORD
Inflammation plays both detrimental and beneficial roles in brain ischemia, depending upon the
timing and severity of the inflammation. Within minutes after injury, injured neurons in the core
and penumbra of the lesion and glial cells in the core produce pro-inflammatory mediators,
cytokines, and reactive oxygen species, which activate both astrocytes and microglia [68].
Activated astrocytes can produce the proinflammatory cytokines IL-6, TNFα, IL-1α and β,
interferon γ, and others [68–70]. High levels of these cytokines can be detrimental to ischemic
recovery [71–75] by directly inducing apoptosis of neuronal cells and/or increasing toxic nitric
oxide levels [76] and inhibiting neurogenesis [77]. Indeed, inactivation of astrocyte NfκB
signaling, shown to induce astrocyte production of pro-inflammatory cytokines [78], decreases
cytokine production and protects neurons after ischemic injury [79]. Hsp72 overexpression is
associated with lower NfκB activation and lower TNFα [80]. In addition to cytokines, reactive
astrocytes also produce chemokines following ischemia [81]. Chemokines upregulate adhesion
molecules in vascular endothelial cells, resulting in attraction of immune cells, which may
worsen ischemia-induced damage [82]. Overall, some aspects of the local inflammatory response
contribute to secondary injury to potentially viable tissue and lead to apoptotic and necrotic
neuronal cell death hours to days after injury [83], while other aspects are beneficial.
Although the potential benefits of inflammation after stroke have received relatively little
attention so far, indirect evidence suggests that some specific inflammatory reactions are
neuroprotective and neuroregenerative [84–91]. In addition to providing defense against the
invasion of pathogens, inflammation is also involved in clearing damaged tissue, and in
angiogenesis, tissue remodeling, and regeneration [89]. This is probably best studied in wound
healing, which is severely compromised if inflammation is inhibited [89, 91]. There is also
evidence suggesting that specific inflammatory factors can be protective in some circumstances.
IL-6, produced by astrocytes acutely after MCAO [69], is likely neuroprotective early after
ischemia [84]. Interestingly, ischemic preconditioning resulting in protection appears to be
dependent on TLR-4 signaling, and is accompanied by increased TNFα, NFκB, and COX-2
expression [90]. Indeed, in vitro work has shown that administration of TNFα in combination
with Hsp70 results in decreased expression of pro-apoptotic proteins following hypoxia [88].
Thus, it is important to consider these factors, along with timing, when trying to determine the
best strategy to reduce damage and improve recovery and regeneration.
5. ASTROCYTE SUPPORT OF NEURONS AFTER STROKE
5.1. Antioxidant Production
One hallmark of the cellular response to ischemia is a rapid, dramatic increase in damaging free
radicals, including nitric oxide (NO), superoxide, and peroxynitrite [92]. Nitric oxide synthetase
levels increase as soon as 10 minutes after induction of MCAO [93], followed by NO production
that persists for at least one week after MCAO [94]. Nitric oxide can cause cell death by
inducing the release of cytochrome-c from mitochondria, leading to apoptosis [95]. Nitric oxide
can also induce necrotic death [96]. Furthermore, the production of nitric oxide and other free
radicals can modify oxidative metabolism and impair ATP production [13, 19]. Changes in
mitochondrial properties can further limit oxidative metabolism [97]. Not surprisingly, several
studies have shown that antioxidant treatment enhances neuroprotection and recovery after
stroke [98–101].
The release of glutathione and SOD by astrocytes has been reported and is suggested to play an
important role in maintaining and enhancing neuronal survival, yet they are able to reduce
ascorbate for further neuronal antioxidant defense Fig. (2) [10, 102–106]. Interestingly, neurons
cocultured with astrocytes exhibit higher levels of glutathione compared with neurons cultured
alone [107]. Although astrocytes upregulate SOD after cerebral ischemia [108], they do not
appear to increase levels of glutathione in ischemic conditions [109]. It is unknown whether
ischemia alters astrocytic ascorbate levels, but osmotic swelling from ischemia results in
increased astrocyte release of ascorbate in vitro [110], suggesting that similar mechanisms may
occur in vivo.
Fig. (2)
Mechanisms of astrocyte support of neurons important in stroke. Antioxidant defense includes
release of glutathione and ascorbate. Regulation of extracellular levels of ions and neuro-
transmitters, especially K+ and glutamate, strongly influences neuronal …
Several treatments that attenuate ischemic injury result in increased glutathione levels [111, 112].
SOD converts superoxide into oxygen and hydrogen peroxide. Similar to glutathione, many
treatments that ameliorate stroke damage are accompanied by an increase in SOD [113, 114].
Furthermore, rodents overexpressing SOD1 have significantly smaller injuries after both focal
and global ischemia [115, 116], while mice with decreased SOD1 have larger infarcts [117].
Finally, ascorbate can also reduce oxidative stress [118]. Treatment with dehydroascorbic acid, a
blood-brain-barrier-permeable precursor to ascorbic acid, is protective after MCAO [119].
Dehydroascorbic acid is taken up by astrocytes and released as ascorbic acid [12], a process
increased by propofol [120], a treatment that can be protective after stroke [121]. In summary,
astrocytes are important producers of antioxidants in the normal CNS, and astrocyte production
of these molecules after stroke may enhance neuronal survival and protect astrocyte function.
5.2. Glutamate Regulation
Astrocytes are key players in the regulation of neuro-transmitters in the CNS. Astrocytes make
glutamine, the precursor for the neurotransmitters glutamate and GABA [122] Fig. (2). Astrocyte
production of neurotransmitter precursors is impaired after MCAO, and alterations in neuro-
transmitter levels occur throughout the brain following stroke, possibly contributing to neuronal
death [123, 124].
Astrocytes are primarily responsible for glutamate uptake in the normal brain using the astrocyte
specific glutamate transporters GLAST and GLT-1 (Fig. 2) [125–127], as excess glutamate leads
to cell death via excitotoxicity [128]. Glutamate transporter levels in astrocytes decrease acutely
following global ischemia [38, 129] and neonatal hypoxia-ischemia [130], most likely
exacerbating neuronal death as a result of glutamate-induced excitoxicity. Despite the therapeutic
potential of increasing astrocyte glutamate transport after stroke, few groups have explored this
possibility. Carnosine, shown to be protective after focal ischemia, may partially be effective
because it prevents loss of GLT-1 on astrocytes, resulting in attenuated excitotoxicity [131]. In a
more direct assessment of how post-ischemic astrocyte glutamate transporters contribute to
neuronal survival, our laboratory has shown that upregulation of GLT-1 on astrocytes using
ceftriaxone protects CA1 neurons after global ischemia [129], similar to its effects in focal
cerebral ischemia [132].
5.3. Potassium Uptake and Energy Metabolism
Astrocytes also regulate neuronal activation by extracellular potassium uptake [133] Fig. (2).
Neurons release potassium after activation, and increased extracellular potassium leads to
neuronal hyperexcitability [133], a phenomenon that occurs in ischemic conditions [134]. In
addition to regulating neuronal activation, proper maintenance of ion gradients, such as
potassium, is important in regulating cell volume in both normal and ischemic conditions
[135, 136]. Astrocytes increase potassium transporter activity in response to transient in
vitro ischemia [137]. Due to its effects on both neuronal activity and cell volume, increasing
astrocytic potassium uptake may be a possible therapeutic target for stroke.
Astrocytes are also integral to normal maintenance of neuronal metabolism. When astrocytes
take up extracellular glutamate as a result of neuronal activity, the Na+/ K+-ATPase, along with
AMPA signaling, triggers astrocyte uptake of glucose from the blood, as astrocytic endfeet
contact capillaries [138, 139]. This glucose is then made into lactate, a substrate for neuronal
energy, to further “fuel” active neurons [140] Fig. (2). As mentioned above, astrocytes produce
glutathione. In addition to its antioxidant properties, glutathione is needed for the conversion of
methylglyoxal, a toxic by-product of metabolism, into D-Lactate by glyoxalase 1 [141].
Although the role of astrocyte metabolism is relatively well-established in normal tissue, the
post-ischemic role of astrocyte metabolism maintenance is less clear [142]. After ischemia,
astrocytes upregulate glucose transporters in order to provide energy to stressed/dying neuronal
cells [143,144]. Ethyl pyruvate, a derivative of the energy substrate pyruvate, is neuroprotective
after stroke only when astrocytes are viable, suggesting that astrocytes are necessary for
improvement in post-ischemic energy metabolism [122].
6. NOVEL STRATEGIES TO IMPROVE THE NEURONAL SUPPORTIVE ROLE OF
ASTROCYTES
Although few studies have specifically targeted astrocytes for repair after stroke, there is some
evidence that this can be a successful strategy. Recent results indicate that induction of BDNF in
astrocytes by galectin-1 reduces neuronal apoptosis in ischemic boundary zone and improves
functional recovery [145]. In addition, protection by pyruvate against glutamate neurotoxicity is
mediated by astrocytes through a glutathione-dependent mechanism [146]. Our recent study
demonstrated that enhancing astrocyte resistance to ischemic stress by overexpressing protective
proteins or antioxidant enzyme results in improved survival of CA1 neurons following forebrain
ischemia Fig. (3) [16]. Two well-studied protective proteins, heat shock protein 72 (Hsp72) and
mitochondrial SOD, were genetically targeted for expression in astrocytes using the astrocyte-
specific human GFAP promoter. In both cases protection was accompanied by preservation of
the astrocytic glutamate transporter GLT-1, and reduced evidence of oxidative stress in the CA1
region [16]. Similarly, selective overexpression of excitatory amino acid transporter 2 (EAAT2)
in astrocytes enhances neuroprotection from moderate hypoxia-ischemia [147].
Fig. (3)
Targeted over-expression of Hsp72 in astrocytes reduces the vulnerability of CA1 neurons to
forebrain ischemia. Selective overexpression of Hsp72 in astrocytes by expressing it from the
astrocyte specific GFAP promoter was achieved by unilateral stereotaxic …
7. TRANSLATING INSIGHTS INTO PROTECTION INTO CLINICAL APPLICATIONS
Many factors have been identified that likely contribute to the failure in translation seen so far
with stroke therapies. Currently, the only approved stroke therapy is thrombolysis induced by
intravenous administration of recombinant tissue plasminogen activator [148]; however, because
of a short therapeutic time window, only a small fraction of patients benefit from this treatment.
Hypothermia is the only accepted acute treatment to reduce brain injury following cardiac arrest
and resuscitation [4]. Thus far many clinical trials have focused on treatments that would likely
be beneficial to neurons, with fewer studies focused on mechanisms that might benefit all cell
types or specifically targeting other cell types, such as astrocytes. Often the consequence of these
treatments on the astrocyte response is not considered. Several examples of past and ongoing
clinical trials are discussed below, with specific attention to how these treatments may alter
astrocyte response or viability.
Several clinical trials have targeted manipulation of the inflammatory response to ischemia, as
stroke patients with systemic inflammation exhibit poorer outcomes [149]. Although anti-
inflammatory therapy decreases infarct size and improves neurological sequelae in experimental
animal models of stroke [150], human trials with anti-neutrophil therapy have not shown a clear
benefit [151, 152]. In addition, recent clinical trials in which anti-CD11/18 antibodies were
administered to human subjects were unsuccessful [153]. Likewise, a double-blinded, placebo-
controlled clinical trial in which anti–ICAM-1 antibody was administered within 6 hours of
stroke symptoms showed disappointing results [151]. In understanding these results it is
important to recall that while experimental stroke is relatively homogeneous concerning size,
territory, and etiology, with more consistent inflammatory response, human stroke is extremely
heterogeneous [154], with different vascular territories and extents of injury. In addition, these
mediators are known to affect many organ systems beyond the central nervous system. Systemic
administration of anti-inflammatory agents may have exacerbated the relative state of
immunocompromise seen in stroke patients, thereby confounding the outcome. Furthermore,
inflammation and astrocyte response are likely closely connected. Although there is little
evidence for a direct relationship between neutrophils and astrocytes, it has been shown that
mice with a blunted inflammatory response exhibit increased loss of GFAP-positive astrocytes
after cortical stab injury [155]. Because astrocytic glial scar formation is important in protection
of spared tissue from further damage [156], it is possible that treatments that drastically attenuate
inflammation lead to a stunted astrocyte response that is deleterious to recovery.
Another drug that has advanced to clinical study is DP-b99, currently in phase III studies for
acute stroke. DP-b99 is a membrane active chelator derivative of the known calcium chelator,
BAPTA spell out [157]. A lipophilic chelator of calcium, zinc and copper ions, DP-b99
sequesters metal ions only within and in to cell membranes. This clinical trial is especially
attractive because sequestration of calcium, zinc, and copper are potentially beneficial not only
to neurons, but also to astrocytes. It has been shown in Alzheimer’s disease that beta amyloid
increases astrocyte calcium influx, which causes decreased glutathione levels [158]. Zinc
chloride is toxic to astrocytes as well as neurons in vitro [159]. Similarly, astrocytes exposed to
neocuprine exhibit increased copper influx and undergo apoptotic cell death [160]. Approaches
that benefit multiple cell types, including astrocytes, are more likely to be successful.
Other current ongoing clinical trials focus on neuroprotective agents for the purpose of aiding
neurological recovery after stroke. Minocycline (Phase I), edavarone (Phase IV), propanolol (a
β-blocker; phase II and III), and more recently arundic acid have been previously shown to be
protective and enhance neuronal survival in stroke [161–165], though by targeting different
mechanisms. Some additional completed and ongoing trials are summarized in Table 1.
Preclinical research needs to consider these clinical results, and assess effects on astrocytes as
well as neurons.
Table 1
Overview of Some Completed and Ongoing Clinical Trials for Stroke
Although anti-inflammatory strategies to diminish ischemic brain injury have failed thus far,
continued elucidation of the complex interactions involved in modulating the inflammatory
response may still enable novel therapeutic approaches that translate successfully into clinical
efficacy.
CONCLUSIONS
Traditionally, stroke research has focused on neurons and often ignored effects on glial cells. It is
increasingly evident that glia are vital to both normal CNS functioning and also play important
roles in neuropathological conditions. Although astrocytes form an inhibitory glial scar following
ischemia, they also perform functions necessary for neuronal survival and well-being, such as
maintaining low extracellular glutamate levels and providing antioxidant protection. Because
they have a great many functions, astrocytes are attractive candidates as therapeutic targets. By
striving to shift astrocytes towards a pro-reparative, neuronal-supportive phenotype following
stroke, future clinical therapies may well be more successful in protecting neurons from ischemic
damage and promoting repair.
ACKNOWLEDGEMENTS
This work was supported by NIH grants CM49831, N5053898, and NS014543 to RGG.
REFERENCES
1. Lloyd-Jones D, Adams RJ, Brown TM, Carnethon M, Dai S, De Simone G, Ferguson TB, Ford E, Furie K,
Gillespie C, Go A, Greenlund K, Haase N, Hailpern S, Ho PM, Howard V, Kissela B, Kittner S, Lackland D,
Lisabeth L, Marelli A, McDermott MM, Meigs J, Mozaffarian D, Mussolino M, Nichol G, Roger VL,
Rosamond W, Sacco R, Sorlie P, Roger VL, Thom T, Wasserthiel-Smoller S, Wong ND, Wylie-Rosett J.
Heart disease and stroke statistics–2010 update: a report from the American Heart
Association.Circulation. 2010;121:e46–e215. [PubMed]
2. Blakeley JO, Llinas RH. Thrombolytic therapy for acute ischemic stroke. J. Neurol.
Sci. 2007;261:55–62.[PubMed]
3. Marder VJ, Jahan R, Gruber T, Goyal A, Arora V. Thrombolysis with plasmin: implications for stroke
treatment. Stroke. 2010;41:S45–S49. [PMC free article] [PubMed]
4. Bernard SA, Gray TW, Buist MD, Jones BM, Silvester W, Gutteridge G, Smith K. Treatment of comatose
survivors of out-of-hospital cardiac arrest with induced hypothermia. N. Engl. J.
Med.2002;346:557–563. [PubMed]
5. Howells DW, Porritt MJ, Rewell SS, O’Collins V, Sena ES, van der Worp HB, Traystman RJ, Macleod MR.
Different strokes for different folks: the rich diversity of animal models of focal cerebral ischemia. J.
Cereb. Blood Flow Metab. 30:1412–1431. [PMC free article] [PubMed]
6. Kettenmann H, Ramson BR. Neuroglia. New York: Oxford University Press; 1995.
7. Nedergaard M, Ransom B, Goldman SA. New roles for astrocytes: redefining the functional
architecture of the brain. Trends Neurosci. 2003;26:523–530. [PubMed]
8. Sofroniew MV, Vinters HV. Astrocytes: biology and pathology. Acta
Neuropathol. 2010;119:7–35.[PMC free article] [PubMed]
9. Goldberg MP, Choi DW. Combined oxygen and glucose deprivation in cortical cell culture: calcium-
dependent and calcium-independent mechanisms of neuronal injury. J.
Neurosci. 1993;13:3510–3524.[PubMed]
10. Swanson RA, Ying W, Kauppinen TM. Astrocyte influences on ischemic neuronal death. Curr. Mol.
Med. 2004;4:193–205. [PubMed]
11. Dringen R, Hirrlinger J. Glutathione pathways in the brain. Biol. Chem. 2003;384:505–516. [PubMed]
12. Wilson JX. Antioxidant defense of the brain: a role for astrocytes. Can. J. Physiol.
Pharmacol.1997;75:1149–1163. [PubMed]
13. Rossi DJ, Brady JD, Mohr C. Astrocyte metabolism and signaling during brain ischemia. Nat.
Neurosci.2007;10:1377–1386. [PubMed]
14. Nedergaard M, Dirnagl U. Role of glial cells in cerebral ischemia. Glia. 2005;50:281–286. [PubMed]
15. Ouyang YB, Voloboueva LA, Xu LJ, Giffard RG. Selective dysfunction of hippocampal CA1 astrocytes
contributes to delayed neuronal damage after transient forebrain ischemia. J.
Neurosci.2007;27:4253–4260. [PMC free article] [PubMed]
16. Xu L, Emery JF, Ouyang YB, Voloboueva LA, Giffard RG. Astrocyte targeted overexpression of Hsp72
or SOD2 reduces neuronal vulnerability to forebrain ischemia. Glia. 2010;58:1042–1049.[PMC free
article] [PubMed]
17. Anderson MF, Blomstrand F, Blomstrand C, Eriksson PS, Nilsson M. Astrocytes and stroke:
networking for survival? Neurochem. Res. 2003;28:293–305. [PubMed]
18. Giffard RG, Swanson RA. Ischemia-induced programmed cell death in
astrocytes. Glia. 2005;50:299–306. [PubMed]
19. Almeida A, Delgado-Esteban M, Bolanos JP, Medina JM. Oxygen and glucose deprivation induces
mitochondrial dysfunction and oxidative stress in neurones but not in astrocytes in primary culture. J.
Neurochem. 2002;81:207–217. [PubMed]
20. Xu L, Sapolsky RM, Giffard RG. Differential sensitivity of murine astrocytes and neurons from
different brain regions to injury. Exp. Neurol. 2001;169:416–424. [PubMed]
21. Zhao G, Flavin MP. Differential sensitivity of rat hippocampal and cortical astrocytes to oxygen-
glucose deprivation injury. Neurosci. Lett. 2000;285:177–180. [PubMed]
22. Lukaszevicz AC, Sampaio N, Guegan C, Benchoua A, Couriaud C, Chevalier E, Sola B, Lacombe P,
Onteniente B. High sensitivity of protoplasmic cortical astroglia to focal ischemia. J. Cereb. Blood Flow
Metab. 2002;22:289–298. [PubMed]
23. Bondarenko A, Chesler M. Rapid astrocyte death induced by transient hypoxia, acidosis, and
extracellular ion shifts. Glia. 2001;34:134–142. [PubMed]
24. Giffard RG, Monyer H, Choi DW. Selective vulnerability of cultured cortical glia to injury by
extracellular acidosis. Brain Res. 1990;530:138–141. [PubMed]
25. Swanson RA, Farrell K, Stein BA. Astrocyte energetics, function, and death under conditions of
incomplete ischemia: a mechanism of glial death in the penumbra. Glia. 1997;21:142–153. [PubMed]
26. Tombaugh GC, Sapolsky RM. Mechanistic distinctions between excitotoxic and acidotic hippocampal
damage in an in vitro model of ischemia. J. Cereb. Blood Flow Metab. 1990;10:527–535. [PubMed]
27. Chen Y, Swanson RA. Astrocytes and brain injury. J. Cereb. Blood Flow
Metab. 2003;23:137–149.[PubMed]
28. Lee MH, Kim H, Kim SS, Lee TH, Lim BV, Chang HK, Jang MH, Shin MC, Shin MS, Kim CJ. Treadmill
exercise suppresses ischemia-induced increment in apoptosis and cell proliferation in hippocampal
dentate gyrus of gerbils. Life Sci. 2003;73:2455–2465. [PubMed]
29. Li Y, Chopp M, Zhang ZG, Zhang RL. Expression of glial fibrillary acidic protein in areas of focal
cerebral ischemia accompanies neuronal expression of 72-kDa heat shock protein. J. Neurol.
Sci.1995;128:134–142. [PubMed]
30. Schroeter M, Schiene K, Kraemer M, Hagemann G, Weigel H, Eysel UT, Witte OW, Stoll G. Astroglial
responses in photochemically induced focal ischemia of the rat cortex. Exp Brain
Res. 1995;106:1–6.[PubMed]
31. Van Beek J, Chan P, Bernaudin M, Petit E, MacKenzie ET, Fontaine M. Glial responses, clusterin, and
complement in permanent focal cerebral ischemia in the mouse. Glia. 2000;31:39–50. [PubMed]
32. Yamashita K, Vogel P, Fritze K, Back T, Hossmann KA, Wiessner C. Monitoring the temporal and
spatial activation pattern of astrocytes in focal cerebral ischemia using in situ hybridization to GFAP
mRNA: comparison with sgp-2 and hsp70 mRNA and the effect of glutamate receptor antagonists. Brain
Res.1996;735:285–297. [PubMed]
33. Liu D, Smith CL, Barone FC, Ellison JA, Lysko PG, Li K, Simpson IA. Astrocytic demise precedes delayed
neuronal death in focal ischemic rat brain. Brain Res. Mol. Brain Res. 1999;68:29–41. [PubMed]
34. Dugan LL, Bruno VM, Amagasu SM, Giffard RG. Glia modulate the response of murine cortical
neurons to excitotoxicity: glia exacerbate AMPA neurotoxicity. J.
Neurosci. 1995;15:4545–4555. [PubMed]
35. Rosenberg PA, Aizenman E. Hundred-fold increase in neuronal vulnerability to glutamate toxicity in
astrocyte-poor cultures of rat cerebral cortex. Neurosci. Lett. 1989;103:162–168. [PubMed]
36. Voloboueva LA, Suh SW, Swanson RA, Giffard RG. Inhibition of mitochondrial function in astrocytes:
implications for neuroprotection. J. Neurochem. 2007;102:1383–1394. [PMC free article] [PubMed]
37. Chen JC, Hsu-Chou H, Lu JL, Chiang YC, Huang HM, Wang HL, Wu T, Liao JJ, Yeh TS. Down-regulation
of the glial glutamate transporter GLT-1 in rat hippocampus and striatum and its modulation by a group
III metabotropic glutamate receptor antagonist following transient global forebrain ischemia. Neuro-
pharmacology. 2005;49:703–714. [PubMed]
38. Yeh TH, Hwang HM, Chen JJ, Wu T, Li AH, Wang HL. Glutamate transporter function of rat
hippocampal astrocytes is impaired following the global ischemia. Neurobiol.
Dis. 2005;18:476–483.[PubMed]
39. Ito U, Hakamata Y, Kawakami E, Oyanagi K. Degeneration of astrocytic processes and their
mitochondria in cerebral cortical regions peripheral to the cortical infarction: heterogeneity of their
disintegration is closely associated with disseminated selective neuronal necrosis and maturation of
injury.Stroke. 2009;40:2173–2181. [PubMed]
40. Ito U, Hakamata Y, Kawakami E, Oyanagi K. Temporary focal cerebral ischemia results in swollen
astrocytic end-feet that compress microvessels and lead to focal cortical infarction. J. Cereb. Blood Flow
Metab. 2010 [PMC free article] [PubMed]
41. Vangeison G, Rempe DA. The Janus-faced effects of hypoxia on astrocyte
function. Neuroscientist.2009;15:579–588. [PMC free article] [PubMed]